ชีวิตของ DNA (กรดดีออกซีไรโบนิวคลีอิก)
คำจำกัดความของ "ดีเอ็นเอ"
ยีน - คือชุดของส่วน DNA ที่กำหนดการก่อตัวของโมเลกุล RNA หรือผลิตภัณฑ์โปรตีน (นักร้อง เอ็ม. เบิร์ก พี. 1998)
มนุษย์มียีนประมาณ 30,000 ยีน ในปริมาตรทั้งหมดของ DNA ยีนโครงสร้าง (เช่น ยีนที่เข้ารหัสโปรตีนที่ใช้ในการสร้างโครงสร้างของร่างกาย) ครอบครองเพียง 3-10%
หน่วยการทำงานที่เล็กที่สุดของ DNA ประกอบด้วยองค์ประกอบต่อไปนี้: ยีนโครงสร้าง, โซนควบคุม, ยีนควบคุม
โครงสร้างของโมเลกุลดีเอ็นเอ
โมเลกุล DNA มีรูปแบบของโพลีเมอร์สายยาวคู่ - โพลีนิวคลีโอไทด์ประกอบด้วยโมโนเมอร์ - นิวคลีโอไทด์ โซ่คู่บิดเป็นเกลียว นี่คือเหตุผลว่าทำไม DNA จึงดูเหมือนบันไดเวียน (ดูภาพด้านบน) นิวคลีโอไทด์แต่ละตัวประกอบด้วยเบสไนโตรเจนหนึ่งในสี่เบส ได้แก่ อะดีนีน (A), กัวนีน (G), ไซโตซีน (C) หรือไทมีน (T), โมเลกุลเพนโตสหนึ่งโมเลกุล (น้ำตาลห้าคาร์บอน) และกรดฟอสฟอริกหนึ่งตกค้าง โดยปกติแล้ว โมเลกุล DNA ประกอบด้วยสายคู่ตรงข้ามสองเส้นที่ก่อตัวเป็นเกลียวคู่ ในกรณีนี้ อะดีนีนของสายหนึ่งจับคู่กับไทมีนของอีกสายหนึ่ง (ทำให้เสถียรด้วยพันธะไฮโดรเจน 2 พันธะ) และกัวนีนมีความเกี่ยวข้องในทำนองเดียวกันกับไซโตซีน (พันธะไฮโดรเจน 3 พันธะ) ลำดับของเบสไนโตรเจนในโมเลกุล DNA จะนำข้อมูลที่จำเป็นสำหรับการสังเคราะห์โปรตีน DNA เป็นโมเลกุลที่ยาวมากประกอบด้วยนิวคลีโอไทด์จำนวนมาก ตัวอย่างเช่น จีโนมของมนุษย์ประกอบด้วยโครโมโซม 46 โครโมโซม ซึ่งมีพื้นฐานมาจากโมเลกุลดีเอ็นเอ ซึ่งประกอบเข้าด้วยกันจากนิวคลีโอไทด์ประมาณ 3 พันล้านคู่
ในยูคาริโอต สารพันธุกรรมจะอยู่ในนิวเคลียสของเซลล์ในโครโมโซม โครโมโซมในสถานะแอคทีฟมีอยู่ในรูปของโครมาติน โครมาตินประกอบด้วย DNA ประมาณ 40%, ฮิสโตน 40% (โปรตีนอัลคาไลน์), โปรตีนโครโมโซมที่ไม่ใช่ฮิสโตนประมาณ 20% และ RNA บางส่วน
วิดีโอ:โครงสร้างโครโมโซม
เราสามารถจำแนก DNA ว่าเป็น “ระบบสิ่งมีชีวิต” หรือ “โมเลกุลที่มีชีวิต” ได้ โดยอ้างว่าสิ่งเหล่านี้เป็นพื้นฐานของชีวิตโดยทั่วไป และยังมีคุณสมบัติที่สำคัญที่สุดหลายประการของชีวิต โดยเฉพาะอย่างยิ่งความสามารถในการสืบพันธุ์ DNA มีความเป็นอิสระและพึ่งพาตนเองได้มากจนสามารถดำรงอยู่ได้แม้จะอยู่นอกเซลล์ - ในรูปแบบของไวรัส ในชีวิตของพวกเขา โมเลกุล DNA ต้องผ่านช่วงชีวิตซึ่งเตือนเราถึงชีวิตของระบบทางชีววิทยาที่ซับซ้อนมากขึ้น นั่นก็คือสิ่งมีชีวิต เหล่านี้คือระยะต่างๆ เช่น การเกิด การเจริญวัย การงาน (กิจกรรม) และ “การตาย”
หัวข้อ: โครงสร้างของดีเอ็นเอ
การบ้าน
- รู้และสามารถเขียนสูตรโครงสร้างของนิวคลีโอไทด์ได้ A, T, G, C, U
- รู้จักโครงสร้างของโมเลกุล DNA และการจัดเรียงของพวกมันเป็นโครโมโซม
- รู้ว่านิวคลีโอไทด์ใน DNA เชื่อมโยงกันในแนวตั้งและแนวนอนอย่างไร แนวคิดของการเชื่อมต่อขนาด 3"-5"
- สามารถใช้ตารางรหัสพันธุกรรมเพื่อสร้างโมเลกุลเปปไทด์ตามส่วน DNA ที่มีขนาดนิวคลีโอไทด์ตั้งแต่ 12 ตัวขึ้นไป
วิดีโอ:โครโมโซม ไมโทซิส การจำลองแบบ
ระยะชีวิตของโมเลกุล DNA
การเกิด (การจำลอง) - การสุก (โครโมโซม) - งาน (การถอดความ) - การควบคุม (การควบคุม) - การเปลี่ยนแปลง (การกลายพันธุ์) - "ความตาย"
1. การจำลองแบบ DNA - การกำเนิดของสาย DNA ลูกสาวใหม่บนสายต้นกำเนิด
2. การสุกของ DNA - การสร้างโครโมโซม
3. การถอดรหัส DNA - งานของ DNA ในรูปแบบของการสังเคราะห์เทมเพลตของ RNA
4. การควบคุมการถอดความ - การควบคุมกิจกรรมการถอดรหัส DNA
5. ซ่อมแซม DNA - ฟื้นฟูพื้นที่ที่เสียหาย
6. การเปลี่ยนแปลงโครงสร้าง DNA - การกลายพันธุ์, transposons
7. การย่อยสลาย DNA - การทำลายล้างในแต่ละรอบการจำลอง
1.การเกิด-การจำลองแบบ
การจำลองดีเอ็นเอเกิดขึ้นง่ายมาก โดยนับ "หนึ่ง สอง สาม" นั่นคือในสามขั้นตอน: 1) การเริ่มต้น 2) การยืดออก 3) การสิ้นสุด
1. การเริ่มต้น - การเริ่มต้น
เป้าหมายเพื่อเริ่มการจำลองแบบ
การจำลองแบบของโมเลกุล DNA ขนาดใหญ่เริ่มต้นด้วยการปรากฏตัวของจุดจำลองแบบ จุดนี้มีลำดับเฉพาะที่มีคู่ A-T มากมาย พื้นที่ดังกล่าวใน DNA เป็นเป้าหมายของโปรตีนที่เริ่มต้นการจำลองแบบอย่างแม่นยำ สำหรับพวกเขาแล้วมีการแนบโปรตีนการรับรู้แบบพิเศษซึ่งช่วยให้มั่นใจได้ถึงการแนบของเอนไซม์การจำลองแบบ เฮลิเคสและ โทพอยโซเมอเรส(ไจราเซส) และเริ่มกระบวนการจำลองแบบ เฮลิเคสคลาย DNA ออกเป็นสองเส้น แบบฟอร์มส้อมการจำลองแบบ โมเลกุล DNA ติดแน่นกับเมทริกซ์นิวเคลียร์ และไม่สามารถหมุนได้อย่างอิสระเมื่อส่วนใดส่วนหนึ่งหลุดออก สิ่งนี้จะบล็อกเฮลิเคสไม่ให้เคลื่อนที่ไปตามโซ่ Topoisomerase จะตัดสาย DNA และลดความตึงเครียดของโครงสร้าง
ที่ทางแยกการจำลองแบบหนึ่งจะมีเฮลิเคสสองตัวที่เคลื่อนที่ไปในทิศทางตรงกันข้าม เส้นที่แยกออกจากกันได้รับการแก้ไขโดยโปรตีนที่จับกับ DNA ไซต์ที่ฟอร์มส้อมการจำลองแบบเรียกว่า "จุดโอริ" (จุดเริ่มต้น - จุดเริ่มต้น) ในยูคาริโอต ส้อมดังกล่าวหลายพันตัวถูกสร้างขึ้นพร้อมกัน ซึ่งทำให้มั่นใจได้ว่ามีอัตราการจำลองสูง
2. การยืดตัว - ความต่อเนื่อง (lengthening)
การเจริญเติบโตของสาย DNA ของลูกสาวบนสายแม่ทั้งสองนั้นเกิดขึ้นแตกต่างกัน DNA polymerase III ของโปรคาริโอตและ δ-หรือ α-DNA polymerase ของยูคาริโอตสามารถสังเคราะห์สาย DNA ใหม่ได้ในทิศทาง 5'>3' เท่านั้น เนื่องจาก สามารถเพิ่มนิวคลีโอไทด์ใหม่ให้กับคาร์บอนที่ตำแหน่ง 3' เท่านั้น แต่ไม่สามารถเติมนิวคลีโอไทด์ที่ตำแหน่ง 5' ได้
เรียกว่าวงจรที่มีทิศทางนี้ ชั้นนำ - การสังเคราะห์สายลูกสาวของ DNA เกิดขึ้นอย่างต่อเนื่อง DNA polymerase III หรือ δ polymerase จะเพิ่มนิวคลีโอไทด์เสริมอย่างต่อเนื่อง
วงจรที่มีขั้ว 3'>5' คือ ล้าหลัง และเสร็จสมบูรณ์เป็นบางส่วน (ในทิศทาง 5’>3’ เช่นกัน) α-DNA polymerase (หรือ DNA polymerase III) สังเคราะห์ส่วนสั้น ๆ บนสายโซ่นี้ - เศษ Okazaki
การสังเคราะห์ชิ้นส่วน Okazaki และห่วงโซ่ชั้นนำเริ่มต้นด้วยการก่อตัว ไพรเมอร์อาร์เอ็นเอ (เมล็ดพันธุ์ ) 10-15 ไรโบนิวคลีโอไทด์ยาวโดยเอนไซม์ ไพรเมส (อาร์เอ็นเอโพลีเมอเรส) DNA polymerases ไม่มีความสามารถในการเริ่มการสังเคราะห์ DNA ตั้งแต่เริ่มต้น แต่สามารถทำได้เท่านั้น สร้างให้เสร็จวงจรที่มีอยู่ ควบคู่ไปกับการก่อตัวของเส้นใยนำหรือชิ้นส่วน Okazaki ไรโบนิวคลีโอไทด์จะถูกลบออกจากไพรเมอร์และแทนที่ด้วยนิวคลีโอไทด์ DNA การแทนที่บริเวณกรดไรโบนิวคลีอิก (ไพรเมอร์) ด้วยบริเวณ DNA เกิดขึ้นโดยใช้ β-DNA polymerase ซึ่งมีฤทธิ์ทั้ง exonuclease และ polymerase
ดังนั้นการจำลองแบบจึงไม่สามารถทำได้หากไม่มีการถอดความชั่วคราวบางส่วน
ความเร็วของการจำลองดีเอ็นเอ (การยืดตัว) อยู่ที่ประมาณ 45,000 นิวคลีโอไทด์ต่อนาที ดังนั้นทางแยกของพ่อแม่จึงคลายตัวด้วยความเร็ว 4,500 รอบต่อนาที ซึ่งเทียบได้กับความเร็วการหมุนของพัดลมระบายความร้อนในคอมพิวเตอร์ (1300-4800 รอบต่อนาที)
3. การยุติ-สิ้นสุด (สิ้นสุด)
การจำลองแบบเสร็จสมบูรณ์เกิดขึ้นเมื่อช่องว่างระหว่างชิ้นส่วน Okazaki เต็มไปด้วยนิวคลีโอไทด์ (โดยมีส่วนร่วมของ DNA ligase) เพื่อสร้าง DNA เส้นคู่ต่อเนื่องกันสองเส้น และเมื่อส้อมการจำลองแบบสองอันมาบรรจบกัน จากนั้น DNA ที่สังเคราะห์ขึ้นจะถูกบิดเป็นเกลียวซุปเปอร์เฮลิกส์
ความถูกต้องของการจำลองนั้นรับประกันได้ด้วยการจับคู่ที่ตรงกันทุกประการของคู่เบสเสริมและการกระทำของ DNA polymerase ซึ่งนอกเหนือจากโพลีเมอเรสแล้ว ยังมีฤทธิ์ของ exonuclease อีกด้วย และสามารถจดจำและแก้ไขข้อผิดพลาดได้ หากมีนิวคลีโอไทด์ที่ไม่ใช่ส่วนประกอบเสริมรวมอยู่ด้วย เอ็นไซม์จะถอยออกไป ตัดมันออก และทำปฏิกิริยาโพลีเมอเรสต่อไป ดังนั้นกระบวนการจำลองแบบจึงมีความแม่นยำสูง
หลังจากการจำลองเสร็จสิ้น DNA methylation จะเกิดขึ้นในบริเวณ –GATC- ที่อะดีนีน (โดยมีการก่อตัวของ N-methyladenine) และสารตกค้างของไซโตซีนด้วยการก่อตัวของ 5-methylcytosine เมทิลเลชั่นไม่รบกวนการเสริมกันของสายโซ่และจำเป็นสำหรับการสร้างโครงสร้างโครโมโซมและการควบคุมการถอดรหัสยีน
ในโปรคาริโอต เช่น แบคทีเรีย DNA สามารถทำซ้ำได้โดยไม่ต้องยืดตัวให้เป็นโมเลกุลเชิงเส้น กล่าวคือ คงเหลืออยู่ในรูปทรงกลมที่มีลักษณะเฉพาะ
วิดีโอ:ป
2. การสุก - การก่อตัวของโครโมโซมและโครมาติน
3. งาน - การถอดความ
วิดีโอ:การปิดกั้นยีน
4. การจัดการ-การควบคุม
5. การบูรณะ (ซ่อมแซม) - การซ่อม
6. การปรับเปลี่ยน-การกลายพันธุ์ .
7. "ความตาย" - การเสื่อมสภาพระหว่างการจำลองแบบ
กรดดีออกซีไรโบนิวคลีอิก หรือ ดีเอ็นเอ เป็นผู้ถ่ายทอดข้อมูลทางพันธุกรรม DNA ส่วนใหญ่ในเซลล์กระจุกตัวอยู่ในนิวเคลียส เป็นองค์ประกอบหลักของโครโมโซม ในยูคาริโอต DNA ยังพบได้ในไมโตคอนเดรียและพลาสติด DNA ประกอบด้วยโมโนนิวคลีโอไทด์ที่เชื่อมต่อกันด้วยโควาเลนต์ซึ่งเป็นตัวแทนของโพลีเมอร์ที่ยาวและไม่มีการแตกแขนง โมโนนิวคลีโอไทด์ที่ประกอบเป็น DNA ประกอบด้วยดีออกซีไรโบส ซึ่งเป็นหนึ่งใน 4 เบสไนโตรเจน (อะดีนีน กวานีน ไซโตซีน และไทมีน) และกรดฟอสฟอริกที่ตกค้าง จำนวนโมโนนิวคลีโอไทด์เหล่านี้มีขนาดใหญ่มาก ตัวอย่างเช่น ในเซลล์โปรคาริโอตที่มีโครโมโซมเดี่ยว DNA เป็นโมเลกุลขนาดใหญ่เดี่ยวที่มีน้ำหนักโมเลกุลมากกว่า 2 x 10 9
โมโนนิวคลีโอไทด์ของสาย DNA หนึ่งสายเชื่อมต่อกันตามลำดับเนื่องจากการก่อตัว พันธะโควาเลนต์ฟอสโฟไดสเตอร์ ระหว่างกลุ่ม OH ของดีออกซีไรโบสของโมโนนิวคลีโอไทด์หนึ่งกับกรดฟอสฟอริกที่ตกค้างของอีกกลุ่มหนึ่ง ด้านหนึ่งของแกนหลักที่เกิดขึ้นของสาย DNA หนึ่งเส้นจะมีเบสไนโตรเจน เปรียบได้กับลูกปัด 4 เม็ดที่ต่างกันบนด้ายเส้นเดียว เพราะ... พวกมันร้อยอยู่บนโซ่น้ำตาลฟอสเฟตเหมือนเดิม
คำถามเกิดขึ้น: สายพอลินิวคลีโอไทด์สายยาวนี้สามารถเข้ารหัสโปรแกรมการพัฒนาของเซลล์หรือแม้แต่สิ่งมีชีวิตทั้งหมดได้อย่างไร คำตอบสำหรับคำถามนี้สามารถหาได้จากการทำความเข้าใจว่าโครงสร้างเชิงพื้นที่ของ DNA เกิดขึ้นได้อย่างไร โครงสร้างของโมเลกุลนี้ถูกถอดรหัสและอธิบายโดย J. Watson และ F. Crick ในปี 1953
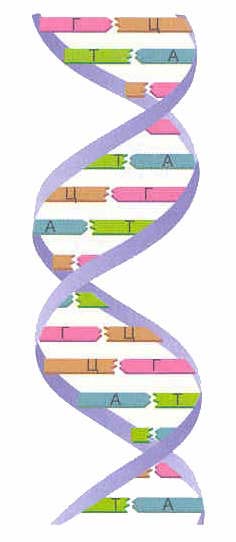
โมเลกุล DNA เป็นสองเส้นที่ขนานกันและก่อตัวขึ้น เกลียวขวา
- ความกว้างของเกลียวนี้อยู่ที่ประมาณ 2 นาโนเมตร แต่ความยาวของมันสามารถเข้าถึงหลายแสนนาโนเมตรได้ วัตสันและคริกได้เสนอแบบจำลองของดีเอ็นเอ โดยฐานดีเอ็นเอทั้งหมดจะอยู่ภายในเกลียว โดยมีแกนหลักที่เป็นน้ำตาล-ฟอสเฟตอยู่ด้านนอก ดังนั้นฐานของห่วงโซ่หนึ่งจึงอยู่ใกล้กับฐานของอีกห่วงโซ่มากที่สุด
ดังนั้นพันธะไฮโดรเจนจึงเกิดขึ้นระหว่างกัน โครงสร้างของเกลียว DNA นั้นสามารถแยกโซ่โพลีนิวคลีโอไทด์ที่ประกอบขึ้นเป็นองค์ประกอบได้หลังจากคลายออกเท่านั้น
เนื่องจากความใกล้ชิดสูงสุดของสาย DNA ทั้งสองสาย องค์ประกอบของมันจึงประกอบด้วยเบสไนโตรเจนประเภทหนึ่ง (อะดีนีนและกัวนีน) และเบสไนโตรเจนประเภทอื่นในปริมาณเท่ากัน (ไทมีนและไซโตซีน) กล่าวคือ สูตรนี้ใช้ได้: A+G=T+C- สิ่งนี้อธิบายได้ด้วยขนาดของฐานไนโตรเจน กล่าวคือ ความยาวของโครงสร้างที่เกิดขึ้นเนื่องจากการก่อตัวของพันธะไฮโดรเจนระหว่างคู่อะดีนีน-ไทมีนและกัวนีน-ไซโตซีนจะอยู่ที่ประมาณ 1.1 นาโนเมตร ขนาดรวมของคู่เหล่านี้สอดคล้องกับขนาดส่วนด้านในของเกลียว DNA ในการสร้างเกลียว คู่ C-T จะมีขนาดเล็กเกินไป และคู่ A-G จะมีขนาดใหญ่เกินไป นั่นคือฐานไนโตรเจนของ DNA สายแรกจะเป็นตัวกำหนดฐานที่อยู่ในตำแหน่งเดียวกันบนสาย DNA อีกเส้นหนึ่ง ความสอดคล้องที่เข้มงวดของนิวคลีโอไทด์ที่อยู่ในโมเลกุล DNA ในสายโซ่คู่ขนานกันเรียกว่า การเสริมกัน (เพิ่มเติม). การทำสำเนาที่แม่นยำหรือ การจำลองแบบ ข้อมูลทางพันธุกรรมเป็นไปได้อย่างแม่นยำด้วยคุณสมบัติของโมเลกุล DNA นี้
ข้อมูลทางชีวภาพถูกเขียนใน DNA ในลักษณะที่สามารถคัดลอกและส่งไปยังเซลล์ลูกหลานได้อย่างแม่นยำ ก่อนที่การแบ่งเซลล์จะเกิดขึ้น การจำลองแบบ (เพิ่มตัวเองเป็นสองเท่า ) ดีเอ็นเอ เนื่องจากแต่ละเกลียวมีลำดับนิวคลีโอไทด์ที่ประกอบกันกับลำดับของนิวคลีโอไทด์คู่ของมัน จริงๆ แล้วพวกมันจึงมีข้อมูลทางพันธุกรรมที่เหมือนกัน หากคุณแยกเกลียวออกและใช้แต่ละเกลียวเป็นแม่แบบ (เทมเพลต) เพื่อสร้างเกลียวที่สอง คุณจะได้เกลียว DNA ใหม่สองเกลียวที่เหมือนกัน นี่เป็นวิธีที่ DNA เพิ่มขึ้นเป็นสองเท่าในเซลล์
เกือบทุกคนเคยได้ยินเกี่ยวกับการมีอยู่ของโมเลกุล DNA ในเซลล์ที่มีชีวิต และรู้ว่าโมเลกุลนี้มีหน้าที่ในการถ่ายทอดข้อมูลทางพันธุกรรม ภาพยนตร์ที่แตกต่างกันจำนวนมาก ไม่ทางใดก็ทางหนึ่ง สร้างขึ้นจากคุณสมบัติของโมเลกุลเล็กๆ แต่น่าภาคภูมิใจและสำคัญมาก
อย่างไรก็ตาม มีเพียงไม่กี่คนที่สามารถอธิบายคร่าวๆ ได้ว่ามีอะไรรวมอยู่ในโมเลกุล DNA บ้าง และกระบวนการอ่านข้อมูลทั้งหมดนี้เกี่ยวกับ "โครงสร้างของสิ่งมีชีวิตทั้งหมด" ทำงานอย่างไร มีเพียงไม่กี่คนที่สามารถอ่าน “กรดดีออกซีไรโบนิวคลีอิก” ได้โดยไม่ลังเลใจ
ลองหาดูว่าโมเลกุลที่สำคัญที่สุดสำหรับเราแต่ละคนประกอบด้วยอะไรและมีลักษณะอย่างไร
โครงสร้างของหน่วยโครงสร้าง - นิวคลีโอไทด์
โมเลกุล DNA ประกอบด้วยหน่วยโครงสร้างหลายหน่วย เนื่องจากเป็นไบโอโพลีเมอร์ โพลีเมอร์เป็นโมเลกุลขนาดใหญ่ที่ประกอบด้วยชิ้นส่วนเล็กๆ จำนวนมากที่เชื่อมต่อกันตามลำดับ เช่นเดียวกับโซ่ที่ประกอบด้วยลิงค์
หน่วยโครงสร้างของโมเลกุลขนาดใหญ่ของ DNA คือนิวคลีโอไทด์ นิวคลีโอไทด์ของโมเลกุล DNA มีสารตกค้างของสารสามชนิด ได้แก่ กรดออร์โธฟอสฟอริก แซ็กคาไรด์ (ดีออกซีไรโบส) และหนึ่งในสี่ฐานที่ประกอบด้วยไนโตรเจนที่เป็นไปได้
โมเลกุล DNA ประกอบด้วยเบสไนโตรเจน: อะดีนีน (A), กัวนีน (G), ไซโตซีน (C) และไทมีน (T)

องค์ประกอบของสายนิวคลีโอไทด์สะท้อนให้เห็นโดยการสลับฐานที่รวมอยู่ในนั้น: -AAGCGTTAGCACGT- ฯลฯ ลำดับสามารถเป็นอะไรก็ได้ นี่คือวิธีที่ DNA สายเดียวเกิดขึ้น
การทำให้เป็นเกลียวของโมเลกุล ปรากฏการณ์ของการเสริมกัน
ขนาดของโมเลกุล DNA ของมนุษย์นั้นใหญ่มาก (แน่นอนว่าขนาดของโมเลกุลอื่น ๆ )! จีโนมของเซลล์เดียว (46 โครโมโซม) มีนิวคลีโอไทด์ประมาณ 3.1 พันล้านคู่ ความยาวของสายโซ่ DNA ที่ประกอบด้วยข้อต่อจำนวนมากนั้นอยู่ที่ประมาณสองเมตร เป็นเรื่องยากที่จะจินตนาการว่าโมเลกุลขนาดใหญ่เช่นนี้สามารถวางไว้ภายในเซลล์เล็กๆ ได้อย่างไร
แต่ธรรมชาติดูแลบรรจุภัณฑ์ที่มีขนาดกะทัดรัดและปกป้องจีโนมของมัน - โซ่ทั้งสองเชื่อมต่อกันด้วยฐานไนโตรเจนและก่อตัวเป็นเกลียวคู่ที่รู้จักกันดี ดังนั้นจึงเป็นไปได้ที่จะลดความยาวของโมเลกุลได้เกือบหกเท่า
ลำดับปฏิสัมพันธ์ของฐานไนโตรเจนนั้นถูกกำหนดโดยปรากฏการณ์ของการเสริมกันอย่างเคร่งครัด อะดีนีนสามารถจับกับไทมีนเท่านั้น ในขณะที่ไซโตซีนสามารถจับกับกัวนีนเท่านั้น คู่ที่เสริมกันเหล่านี้ประกอบเข้าด้วยกันเหมือนกุญแจและแม่กุญแจ เหมือนชิ้นส่วนปริศนา
ทีนี้ลองคำนวณจำนวนหน่วยความจำในคอมพิวเตอร์ (หรือบนแฟลชไดรฟ์) ข้อมูลทั้งหมดเกี่ยวกับโมเลกุลขนาดเล็ก (ในระดับโลกของเรา) นี้ที่ควรใช้ จำนวนคู่นิวคลีโอไทด์คือ 3.1x10 9 มีทั้งหมด 4 ค่า ซึ่งหมายความว่าข้อมูล 2 บิต (2 2 ค่า) ก็เพียงพอสำหรับหนึ่งคู่ เราคูณทั้งหมดนี้เข้าด้วยกันแล้วได้ 6200000000 บิต หรือ 775000000 ไบต์ หรือ 775000 กิโลไบต์ หรือ 775 เมกะไบต์ ซึ่งใกล้เคียงกับความจุของซีดีหรือปริมาณของตอนภาพยนตร์ความยาว 40 นาทีในคุณภาพเฉลี่ย
การสร้างโครโมโซม ความมุ่งมั่นของจีโนมมนุษย์
นอกเหนือจากการทำให้เป็นเกลียวแล้ว โมเลกุลยังถูกบดอัดซ้ำหลายครั้ง เกลียวคู่เริ่มบิดเหมือนลูกบอลด้าย - กระบวนการนี้เรียกว่าซูเปอร์คอยล์และเกิดขึ้นด้วยความช่วยเหลือของโปรตีนฮิสโตนพิเศษซึ่งโซ่นั้นพันเหมือนแกนม้วน
กระบวนการนี้ทำให้ความยาวของโมเลกุลสั้นลงอีก 25-30 เท่า จากการบรรจุภัณฑ์อีกหลายระดับ โมเลกุล DNA หนึ่งโมเลกุลที่มีโปรตีนเสริมจึงกลายเป็นโครโมโซมที่มีขนาดกะทัดรัดมากขึ้นเรื่อยๆ

ข้อมูลทั้งหมดที่เกี่ยวข้องกับรูปร่าง ลักษณะ และการทำงานของร่างกายของเราถูกกำหนดโดยชุดของยีน ยีนเป็นส่วนที่กำหนดไว้อย่างเคร่งครัดของโมเลกุล DNA ประกอบด้วยลำดับนิวคลีโอไทด์ที่ไม่เปลี่ยนแปลง นอกจากนี้ ยีนยังถูกกำหนดอย่างเคร่งครัดไม่เพียงแต่จากองค์ประกอบของมันเท่านั้น แต่ยังรวมถึงตำแหน่งของยีนที่สัมพันธ์กับส่วนอื่นๆ ของสายโซ่ด้วย
กรดริโบนิวคลีอิกและบทบาทในการสังเคราะห์โปรตีน
นอกจาก DNA แล้วยังมีกรดนิวคลีอิกประเภทอื่น ๆ - เมทริกซ์, การถ่ายโอนและไรโบโซมอล RNA (กรดไรโบนิวคลีอิก) สายโซ่ RNA นั้นเล็กกว่าและสั้นกว่ามาก จึงสามารถทะลุผ่านเยื่อหุ้มนิวเคลียสได้
โมเลกุล RNA ก็เป็นไบโอโพลีเมอร์เช่นกัน ชิ้นส่วนที่มีโครงสร้างคล้ายกับชิ้นส่วนที่ประกอบเป็น DNA ยกเว้นแซ็กคาไรด์เล็กน้อย (น้ำตาลแทนดีออกซีไรโบส) เบสไนโตรเจนมีสี่ประเภท: A, G, C ที่คุ้นเคยและยูราซิล (U) แทนไทมีน ภาพด้านบนแสดงให้เห็นทั้งหมดนี้อย่างชัดเจน
โมเลกุลขนาดใหญ่ของ DNA สามารถส่งข้อมูลไปยัง RNA ในรูปแบบที่คลายออกได้ การคลายเกลียวเกิดขึ้นด้วยความช่วยเหลือของเอนไซม์พิเศษซึ่งจะแยกเกลียวคู่ออกเป็นสายโซ่ที่แยกจากกัน - เหมือนครึ่งหนึ่งของซิปที่แยกออกจากกัน
ในเวลาเดียวกัน ขนานกับสาย DNA ก็มีการสร้างสายโซ่ RNA เสริมขึ้นมา หลังจากคัดลอกข้อมูลและเข้าสู่สภาพแวดล้อมของเซลล์จากนิวเคลียสแล้ว สายโซ่ RNA จะเริ่มกระบวนการสังเคราะห์โปรตีนที่เข้ารหัสโดยยีน การสังเคราะห์โปรตีนเกิดขึ้นในออร์แกเนลล์เซลล์พิเศษ - ไรโบโซม
ขณะที่ไรโบโซมอ่านสายโซ่ จะเป็นตัวกำหนดว่ากรดอะมิโนจำเป็นต้องเชื่อมต่อกันตามลำดับใด ขณะอ่านข้อมูลลงใน RNA จากนั้นสายโซ่กรดอะมิโนที่สังเคราะห์ขึ้นจะมีรูปทรง 3 มิติที่เฉพาะเจาะจง
โมเลกุลโครงสร้างขนาดใหญ่นี้เป็นโปรตีนที่สามารถทำหน้าที่เข้ารหัสของเอนไซม์ ฮอร์โมน ตัวรับ และวัสดุก่อสร้างได้
ข้อสรุป
สำหรับสิ่งมีชีวิตใดๆ โปรตีนคือผลิตภัณฑ์ขั้นสุดท้ายของแต่ละยีน เป็นโปรตีนที่กำหนดความหลากหลายของรูปแบบ คุณสมบัติ และคุณภาพที่ถูกเข้ารหัสในเซลล์ของเรา
เรียนผู้อ่านบล็อก คุณรู้ไหมว่า DNA อยู่ที่ไหน แสดงความคิดเห็นหรือแสดงความคิดเห็นเกี่ยวกับสิ่งที่คุณอยากรู้ นี่จะมีประโยชน์มากสำหรับใครบางคน!
หลักการของการถ่ายทอดทางพันธุกรรมได้รับการระบุครั้งแรกในคริสต์ทศวรรษ 1900 เมื่อหลักการทางธรรมชาติได้รับการพัฒนาและนำแนวคิดเกี่ยวกับจีโนมมนุษย์และยีนโดยเฉพาะมาใช้ (พร้อมคำจำกัดความที่สมบูรณ์) การวิจัยของพวกเขาช่วยให้นักวิทยาศาสตร์ค้นพบความลับของการถ่ายทอดทางพันธุกรรม และกลายเป็นแรงผลักดันในการศึกษานี้ โรคทางพันธุกรรมและธรรมชาติของพวกเขา
จีโนมมนุษย์: แนวคิดทั่วไป
เพื่อทำความเข้าใจว่ายีนคืออะไรและกระบวนการสืบทอดโดยสิ่งมีชีวิตที่มีคุณสมบัติและคุณสมบัติบางอย่าง คุณควรรู้และเข้าใจข้อกำหนดและข้อกำหนดพื้นฐาน การสรุปแนวคิดพื้นฐานโดยย่อจะให้โอกาสในการเจาะลึกในหัวข้อนี้
ยีนของมนุษย์เป็นส่วนหนึ่งของสายโซ่ (กรดดีออกซีไรโบนิวคลีอิกในรูปของโมเลกุลขนาดใหญ่) ที่กำหนดลำดับของโพลีเปปไทด์บางชนิด (ตระกูลของกรดอะมิโน) และ มีข้อมูลทางพันธุกรรมขั้นพื้นฐานจากพ่อแม่ถึงลูก
พูดง่ายๆ ก็คือ ยีนบางตัวมีข้อมูลเกี่ยวกับโครงสร้างของโปรตีนและถ่ายทอดจากพ่อแม่สู่ลูก โดยทำซ้ำโครงสร้างของโพลีเปปไทด์และถ่ายทอดทางพันธุกรรม
จีโนมมนุษย์เป็นแนวคิดทั่วไปที่แสดงถึงยีนจำเพาะจำนวนหนึ่ง ได้รับการแนะนำครั้งแรกโดย Hans Winkler ในปี 1920 แต่เมื่อเวลาผ่านไป ความหมายดั้งเดิมของคำนี้ก็เปลี่ยนไปบ้าง
ในตอนแรกมันกำหนดจำนวนโครโมโซมจำนวนหนึ่ง (ไม่จับคู่และเป็นเดี่ยว) และหลังจากนั้นไม่นานปรากฎว่าจีโนมมีโครโมโซมที่จับคู่ 23 โครโมโซมและกรดไมโตคอนเดรียลดีออกซีไรโบนิวคลีอิก
ข้อมูลทางพันธุกรรมคือข้อมูลที่มีอยู่ใน DNA และเรียงลำดับการสร้างโปรตีนในรูปแบบของรหัสนิวคลีโอไทด์ นอกจากนี้ยังควรกล่าวถึงด้วยว่าข้อมูลประเภทนี้พบได้ทั้งภายในและภายนอกขอบเขต
 ยีนของมนุษย์ได้รับการศึกษามานานหลายปี ในระหว่างนั้นยีนดังกล่าวก็มีชีวิตขึ้นมา การทดลองมากมาย- ยังคงมีการทดลองเพื่อให้ข้อมูลใหม่แก่นักวิทยาศาสตร์
ยีนของมนุษย์ได้รับการศึกษามานานหลายปี ในระหว่างนั้นยีนดังกล่าวก็มีชีวิตขึ้นมา การทดลองมากมาย- ยังคงมีการทดลองเพื่อให้ข้อมูลใหม่แก่นักวิทยาศาสตร์
การวิจัยเมื่อเร็วๆ นี้แสดงให้เห็นชัดเจนว่ากรดดีออกซีไรโบนิวคลีอิกไม่ได้มีโครงสร้างที่ชัดเจนและสม่ำเสมอเสมอไป
มีสิ่งที่เรียกว่ายีนไม่ต่อเนื่อง การเชื่อมต่อของยีนถูกขัดจังหวะ ซึ่งทำให้ทฤษฎีก่อนหน้านี้ทั้งหมดเกี่ยวกับความคงตัวของอนุภาคเหล่านี้ไม่ถูกต้อง การเปลี่ยนแปลงเกิดขึ้นเป็นครั้งคราวซึ่งนำมาซึ่งการเปลี่ยนแปลงโครงสร้างของกรดดีออกซีไรโบนิวคลีอิก
ประวัติความเป็นมาของการค้นพบ
คำศัพท์ทางวิทยาศาสตร์ถูกกำหนดครั้งแรกในปี 1909 โดยนักวิทยาศาสตร์ Vilhelm Johansen ซึ่งเป็นนักพฤกษศาสตร์ที่โดดเด่นในเดนมาร์ก
สำคัญ!ในปี พ.ศ. 2455 คำว่า "พันธุศาสตร์" ปรากฏขึ้นซึ่งต่อมาได้กลายเป็นชื่อของแผนกทั้งหมด พระองค์คือผู้ที่ศึกษายีนของมนุษย์
การวิจัยอนุภาคได้เริ่มต้นขึ้นแล้ว ยาวนานก่อนศตวรรษที่ 20(ไม่มีข้อมูลปีที่แน่นอน) และประกอบด้วยหลายขั้นตอน:
- ในปี พ.ศ. 2411 ดาร์วินนักวิทยาศาสตร์ชื่อดังได้เสนอสมมติฐานเรื่องการเกิดตับ ในนั้นเขาบรรยายถึงการแยกตัวของอัญมณี ดาร์วินเชื่อว่าเจมมูลคือส่วนเฉพาะของเซลล์ที่เซลล์เพศได้ถูกสร้างขึ้น
- ไม่กี่ปีต่อมา อูโก เดอ วรีส์ได้ก่อตั้งทฤษฎีของตัวเองขึ้นมา ซึ่งแตกต่างไปจากทฤษฎีของดาร์วิน ซึ่งเขาบรรยายถึงกระบวนการของการสร้างเนื้อร้ายภายในเซลล์ เขาเชื่อว่ามีอนุภาคอยู่ในทุกเซลล์ และมีหน้าที่รับผิดชอบต่อคุณสมบัติบางอย่างของการสืบทอดสายพันธุ์ เขาเรียกอนุภาคเหล่านี้ว่า "แพนยีน" ความแตกต่างระหว่างสองสมมติฐานคือดาร์วินถือว่าเจมมูลเป็นส่วนหนึ่งของเนื้อเยื่อและอวัยวะภายใน โดยไม่คำนึงถึงประเภทของสัตว์ และเดอ ไวรีส์ได้นำเสนอตับจีนของเขาเป็นสัญญาณของการถ่ายทอดทางพันธุกรรมภายในสปีชีส์หนึ่งๆ
- W. Johansen ในปี 1900 ให้นิยามปัจจัยทางพันธุกรรมว่าเป็นยีน โดยนำส่วนที่สองมาจากคำที่เดอไวรีส์ใช้ เขาใช้คำนี้ให้คำจำกัดความว่า "เชื้อโรค" ซึ่งเป็นอนุภาคที่เป็นกรรมพันธุ์ ในเวลาเดียวกัน นักวิทยาศาสตร์ได้เน้นย้ำถึงความเป็นอิสระของคำนี้จากทฤษฎีที่หยิบยกมาก่อนหน้านี้
นักชีววิทยาและนักสัตววิทยาได้ศึกษาปัจจัยทางพันธุกรรมมาเป็นเวลานาน แต่ตั้งแต่ต้นศตวรรษที่ 20 เท่านั้นที่พันธุกรรมเริ่มพัฒนาอย่างรวดเร็วเผยให้เห็นความลับของการสืบทอดต่อผู้คน
การถอดรหัสจีโนมมนุษย์
นับตั้งแต่วินาทีที่นักวิทยาศาสตร์ค้นพบการมีอยู่ของยีนในร่างกายมนุษย์ พวกเขาก็เริ่มตรวจสอบคำถามเกี่ยวกับข้อมูลที่มีอยู่ในนั้น เป็นเวลากว่า 80 ปีที่นักวิทยาศาสตร์พยายามถอดรหัสมัน จนถึงปัจจุบันพวกเขาประสบความสำเร็จอย่างมากในเรื่องนี้ซึ่งมอบให้ โอกาสในการมีอิทธิพลเกี่ยวกับกระบวนการทางพันธุกรรมและการเปลี่ยนแปลงโครงสร้างของเซลล์ในรุ่นต่อไป
ประวัติความเป็นมาของการถอดรหัส DNA ประกอบด้วยช่วงเวลาที่กำหนดหลายประการ:
- ศตวรรษที่ 19 - จุดเริ่มต้นของการศึกษากรดนิวคลีอิก
- 1868 - F. Miescher แยกนิวเคลียสหรือ DNA ออกจากเซลล์เป็นครั้งแรก
- ในช่วงกลางศตวรรษที่ 20 O. Avery และ F. Griffith ค้นพบโดยใช้การทดลองกับหนูว่าเป็นกรดนิวคลีอิกที่รับผิดชอบกระบวนการเปลี่ยนแปลงของแบคทีเรีย
- บุคคลแรกที่แสดง DNA ให้โลกเห็นคืออาร์. แฟรงคลิน หลายปีหลังจากการค้นพบกรดนิวคลีอิก เขาได้ถ่ายภาพ DNA แบบสุ่มโดยใช้รังสีเอกซ์ขณะตรวจดูโครงสร้างของผลึก
- ในปี พ.ศ. 2496 ได้มีการให้คำจำกัดความที่ชัดเจนเกี่ยวกับหลักการของการสืบพันธุ์ของสิ่งมีชีวิตในทุกสายพันธุ์
ความสนใจ! นับตั้งแต่ DNA double helix เปิดตัวสู่สาธารณะเป็นครั้งแรก จึงมีการค้นพบมากมายที่ให้ข้อมูลเชิงลึกเกี่ยวกับธรรมชาติของ DNA และวิธีการทำงาน
 โดยผู้ชาย ผู้ค้นพบยีนนี้ Gregor Mendel ถือเป็นกลุ่มแรกที่ค้นพบรูปแบบบางอย่างในสายโซ่ทางพันธุกรรม
โดยผู้ชาย ผู้ค้นพบยีนนี้ Gregor Mendel ถือเป็นกลุ่มแรกที่ค้นพบรูปแบบบางอย่างในสายโซ่ทางพันธุกรรม
แต่การถอดรหัส DNA ของมนุษย์นั้นขึ้นอยู่กับการค้นพบของนักวิทยาศาสตร์อีกคนหนึ่งคือ Frederick Sanger ซึ่งเป็นผู้พัฒนาวิธีการอ่านลำดับกรดอะมิโนโปรตีนและลำดับการสร้าง DNA เอง
ต้องขอบคุณผลงานของนักวิทยาศาสตร์หลายคนในช่วงสามศตวรรษที่ผ่านมา กระบวนการก่อตัว ลักษณะ และจำนวนยีนในจีโนมมนุษย์ได้รับการชี้แจงให้ชัดเจนขึ้น
ในปี 1990 ได้เริ่มต้นขึ้น โครงการระหว่างประเทศ The Human Genome กำกับโดย เจมส์ วัตสัน เป้าหมายของเขาคือค้นหาว่านิวคลีโอไทด์ใน DNA ถูกจัดเรียงตามลำดับใด และเพื่อระบุยีนประมาณ 25,000 ยีนในมนุษย์ ต้องขอบคุณโปรเจ็กต์นี้ บุคคลควรจะได้รับความเข้าใจอย่างถ่องแท้เกี่ยวกับการก่อตัวของ DNA และตำแหน่งของชิ้นส่วนที่เป็นส่วนประกอบทั้งหมดตลอดจนกลไกการสร้างยีน
ควรชี้แจงให้ชัดเจนว่าโปรแกรมไม่ได้กำหนดไว้เพื่อกำหนดลำดับกรดนิวคลีอิกทั้งหมดในเซลล์ แต่มีเพียงบางพื้นที่เท่านั้น เริ่มต้นในปี 1990 แต่เพียงในปี 2000 เท่านั้นที่ได้มีการเผยแพร่ร่างผลงานและการศึกษาฉบับเต็ม แล้วเสร็จ - ในปี พ.ศ. 2546- การวิจัยตามลำดับยังคงดำเนินต่อไป และ 8% ของบริเวณเฮเทอโรโครมาติกยังไม่สามารถระบุได้
เป้าหมายและวัตถุประสงค์
เช่นเดียวกับโครงการทางวิทยาศาสตร์อื่นๆ จีโนมมนุษย์กำหนดเป้าหมายและวัตถุประสงค์เฉพาะของตัวเอง ในตอนแรก นักวิทยาศาสตร์ตั้งใจที่จะระบุลำดับของนิวคลีโอไทด์ตั้งแต่ 3 พันล้านตัวขึ้นไป จากนั้น กลุ่มนักวิจัยที่แยกจากกันแสดงความปรารถนาที่จะกำหนดลำดับของโพลีเมอร์ชีวภาพไปพร้อมๆ กัน ซึ่งอาจเป็นกรดอะมิโนหรือนิวคลีโอไทด์ ส่งผลให้ เป้าหมายหลักของโครงการดูเหมือนว่านี้:
- สร้างแผนที่จีโนม
- สร้างแผนที่โครโมโซมของมนุษย์
- ระบุลำดับการก่อตัวของโพลีเปปไทด์
- สร้างวิธีการจัดเก็บและวิเคราะห์ข้อมูลที่รวบรวม
- สร้างเทคโนโลยีที่จะช่วยให้บรรลุเป้าหมายข้างต้นทั้งหมด
รายการงานนี้ยังขาดงานที่สำคัญพอๆ กัน แต่ไม่ชัดเจนนัก นั่นคือการศึกษาผลที่ตามมาด้านจริยธรรม กฎหมาย และสังคมของการวิจัยดังกล่าว ปัญหาเกี่ยวกับพันธุกรรมอาจทำให้เกิดความขัดแย้งในหมู่ผู้คนและนำไปสู่ความขัดแย้งที่ร้ายแรง ดังนั้น นักวิทยาศาสตร์จึงตั้งเป้าหมายที่จะค้นพบแนวทางแก้ไขสำหรับความขัดแย้งเหล่านี้ก่อนที่จะเกิดขึ้น
ความสำเร็จ
 ลำดับทางพันธุกรรมได้แก่ ปรากฏการณ์ที่ไม่เหมือนใครซึ่งสังเกตได้ในร่างกายของทุกคนในรูปแบบใดรูปแบบหนึ่ง
ลำดับทางพันธุกรรมได้แก่ ปรากฏการณ์ที่ไม่เหมือนใครซึ่งสังเกตได้ในร่างกายของทุกคนในรูปแบบใดรูปแบบหนึ่ง
โครงการบรรลุเป้าหมายทั้งหมดเร็วกว่าที่นักวิจัยคาดไว้ เมื่อสิ้นสุดโครงการ พวกเขาถอดรหัส DNA ได้ประมาณ 99.99% แม้ว่านักวิทยาศาสตร์จะมอบหมายหน้าที่ในการเรียงลำดับข้อมูลเพียง 95% เท่านั้น . ปัจจุบันนี้แม้โครงการจะประสบความสำเร็จ แต่ก็ยังมี พื้นที่ที่ยังไม่ได้สำรวจกรดดีออกซีไรโบนิวคลีอิก
จากผลการวิจัยพบว่ามียีนจำนวนเท่าใดในร่างกายมนุษย์ (ประมาณ 20-25,000 ยีนในจีโนม) และทั้งหมดมีลักษณะเฉพาะ:
- ปริมาณ;
- ที่ตั้ง;
- คุณสมบัติโครงสร้างและการใช้งาน
จีโนมมนุษย์--การวิจัย การถอดรหัส
การถอดรหัสจีโนมมนุษย์
บทสรุป
ข้อมูลทั้งหมดจะถูกนำเสนอโดยละเอียดในแผนที่พันธุกรรมของร่างกายมนุษย์ การดำเนินโครงการวิทยาศาสตร์ที่ซับซ้อนดังกล่าวไม่เพียงแต่ให้ความรู้ทางทฤษฎีจำนวนมหาศาลสำหรับวิทยาศาสตร์พื้นฐานเท่านั้น แต่ยังมีผลกระทบอย่างไม่น่าเชื่อต่อความเข้าใจเกี่ยวกับพันธุกรรมอีกด้วย ในทางกลับกันสิ่งนี้ไม่สามารถส่งผลกระทบต่อกระบวนการป้องกันและรักษาโรคทางพันธุกรรมได้
การค้นพบของนักวิทยาศาสตร์ช่วยเร่งการวิจัยระดับโมเลกุลอื่นๆ และมีส่วนช่วยในเรื่องนี้ การค้นหาพื้นฐานทางพันธุกรรมอย่างมีประสิทธิภาพในโรคที่สืบทอดมาและจูงใจต่อพวกเขา ผลลัพธ์สามารถมีอิทธิพลต่อการค้นพบยาที่เหมาะสมสำหรับการป้องกันโรคต่างๆ เช่น โรคหลอดเลือด หัวใจขาดเลือด อาการป่วยทางจิต และมะเร็ง
(พร้อมกับ RNA) ซึ่งเป็นโพลีเมอร์หรือที่แม่นยำกว่านั้นคือโพลีนิวคลีโอไทด์ (โมโนเมอร์ - นิวคลีโอไทด์)
DNA มีหน้าที่จัดเก็บและส่งรหัสพันธุกรรมระหว่างการแบ่งเซลล์ โดยอาศัยโมเลกุล DNA ที่ทำให้ทราบถึงพันธุกรรมและความแปรปรวน RNA ทุกประเภทถูกสังเคราะห์จาก DNA นอกจากนี้ RNA ประเภทต่างๆ ยังร่วมกันรับประกันการสังเคราะห์โปรตีนของเซลล์ เช่น พวกมันใช้ข้อมูลทางพันธุกรรม
ในเซลล์ยูคาริโอต DNA ส่วนใหญ่อยู่ในนิวเคลียส ซึ่งมันไปรวมตัวกันด้วยโปรตีนชนิดพิเศษเพื่อสร้างโครโมโซม ในเซลล์โปรคาริโอตจะมีโมเลกุล DNA ทรงกลมขนาดใหญ่ (หรือเชิงเส้น) หนึ่งโมเลกุล (และซับซ้อนด้วยโปรตีนด้วย) นอกจากนี้เซลล์ยูคาริโอตยังมี DNA ของตัวเองในไมโตคอนเดรียและคลอโรพลาสต์
ในกรณีของ DNA นิวคลีโอไทด์แต่ละตัวประกอบด้วย 1) ฐานไนโตรเจนซึ่งอาจเป็นอะดีนีน กวานีน ไซโตซีน หรือไทมีน 2) ดีออกซีไรโบส 3) กรดฟอสฟอริก
ลำดับของนิวคลีโอไทด์ในสายโซ่ DNA จะกำหนดโครงสร้างปฐมภูมิของโมเลกุล DNA มีลักษณะเป็นโครงสร้างรองของโมเลกุลในรูปของเกลียวคู่ (ส่วนใหญ่มักจะถนัดขวา) ในกรณีนี้ DNA สองเส้นเชื่อมต่อถึงกันด้วยพันธะไฮโดรเจนที่เกิดขึ้นระหว่างฐานไนโตรเจนเสริม
อะดีนีนเป็นส่วนเสริมของไทมีน และกัวนีนเป็นส่วนเสริมของไซโตซีน พันธะไฮโดรเจนสองอันเกิดขึ้นระหว่างอะดีนีนและไทมีน และอีกสามพันธะระหว่างกัวนีนกับไซโตซีน ดังนั้น guanine และ cytosine จึงเชื่อมต่อกันแน่นขึ้นเล็กน้อย (แม้ว่าหลักการแล้วพันธะไฮโดรเจนจะอ่อนแอก็ตาม) จำนวนพันธะถูกกำหนดโดยลักษณะโครงสร้างของโมเลกุล
อะดีนีนและกัวนีนเป็นพิวรีนและประกอบด้วยวงแหวนสองวง ไทมีนและไซโตซีนเป็นฐานไพริมิดีนแบบวงแหวนเดี่ยว ดังนั้น ระหว่างแกนหลัก (ประกอบด้วยดีออกซีไรโบสและกรดฟอสฟอริกสลับกัน) ของสาย DNA สองสาย สำหรับนิวคลีโอไทด์คู่ใด ๆ ที่มีสายโซ่ต่างกันก็จะมีวงแหวนสามวงเสมอ (เนื่องจากพิวรีนสองวงจะประกอบกันเฉพาะกับวงแหวนเดี่ยวบางวงเท่านั้น ไพริมิดีน) ซึ่งช่วยให้ความกว้างระหว่างสายของโมเลกุล DNA คงเดิมตลอด (ประมาณ 2.3 นาโนเมตร)
มีนิวคลีโอไทด์ประมาณ 10 ตัวในหนึ่งรอบของเกลียว ความยาวของนิวคลีโอไทด์หนึ่งตัวจะอยู่ที่ประมาณ 0.34 นาโนเมตร ความยาวของโมเลกุล DNA มักจะใหญ่โตเกินกว่านิวคลีโอไทด์หลายล้านตัว ดังนั้น เพื่อให้พอดีกับนิวเคลียสของเซลล์ให้แน่นยิ่งขึ้น DNA จึงต้องเผชิญกับ "คอยล์ยิ่งยวด" ในระดับที่แตกต่างกัน
เมื่ออ่านข้อมูลจาก DNA (นั่นคือการสังเคราะห์ RNA ในกระบวนการนี้เรียกว่า การถอดเสียง) despiralization เกิดขึ้น (กระบวนการย้อนกลับของเกลียว) โซ่ทั้งสองแยกออกจากกันภายใต้การกระทำของเอนไซม์พิเศษ พันธะไฮโดรเจนมีความอ่อนแอ ดังนั้นการแยกและการเชื่อมโยงโซ่ตามมาจึงเกิดขึ้นโดยใช้พลังงานเพียงเล็กน้อย RNA ถูกสังเคราะห์จาก DNA ตามหลักการเดียวกันของการเสริมกัน แทนที่จะเป็นไทมีนใน RNA เท่านั้น ยูราซิลจะเสริมกับอะดีนีน
รหัสพันธุกรรมที่เขียนบนโมเลกุล DNA ประกอบด้วยแฝดสาม (ลำดับของนิวคลีโอไทด์สามตัว) ซึ่งเป็นตัวแทนของกรดอะมิโนหนึ่งตัว (โปรตีนโมโนเมอร์) อย่างไรก็ตาม DNA ส่วนใหญ่ไม่ได้เข้ารหัสโปรตีน ความสำคัญของส่วนต่างๆ ของโมเลกุลนั้นแตกต่างกันไปและส่วนใหญ่ยังไม่เป็นที่เข้าใจทั้งหมด
ก่อนที่เซลล์จะแบ่งตัว ปริมาณ DNA จะเพิ่มขึ้นสองเท่าเสมอ กระบวนการนี้เรียกว่า การจำลองแบบ- มันเป็นลักษณะกึ่งอนุรักษ์นิยม: สายโซ่ของโมเลกุล DNA หนึ่งโมเลกุลจะแยกจากกัน และแต่ละโมเลกุลจะเสร็จสมบูรณ์ด้วยสายโซ่เสริมใหม่ของตัวเอง เป็นผลให้จากโมเลกุล DNA ที่มีเกลียวคู่หนึ่งโมเลกุลจะได้รับ DNA ที่มีเกลียวคู่สองอันซึ่งเหมือนกับโมเลกุลตัวแรก
ใน DNA สายพอลินิวคลีโอไทด์มีหลายทิศทาง กล่าวคือ โดยที่สายหนึ่งมีปลาย 5 นิ้ว (กรดฟอสฟอริกติดอยู่กับอะตอมคาร์บอนที่ 5 ของดีออกซีไรโบส) อีกสายหนึ่งมีปลาย 3 นิ้ว (คาร์บอนปราศจากกรดฟอสฟอริก)
ในระหว่างการจำลองและการถอดรหัส การสังเคราะห์จะดำเนินการในทิศทางจากปลาย 5 นิ้วไปยังปลาย 3 นิ้วเสมอ เนื่องจากนิวคลีโอไทด์ใหม่สามารถเกาะติดกับอะตอมคาร์บอนอิสระขนาด 3 นิ้วเท่านั้น
โครงสร้างและบทบาทของ DNA ในฐานะสารที่รับผิดชอบต่อข้อมูลทางพันธุกรรมได้รับการชี้แจงในช่วงทศวรรษที่ 40-50 ของศตวรรษที่ 20 ในปี พ.ศ. 2496 ดี. วัตสัน และ เอฟ. คริก ได้ค้นพบโครงสร้างดีเอ็นเอแบบเกลียวคู่ ก่อนหน้านี้ E. Chargaff พบว่าใน DNA ปริมาณไทมีนจะสัมพันธ์กับอะดีนีนเสมอ และปริมาณกัวนีนต่อไซโตซีน